






ORIGIN AND RELATIONSHIP
Possible systematic positions of birds. Redrawn from Padian et al (1998).
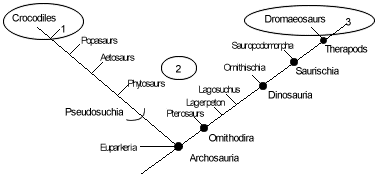
The Thecodont Hypothesis
The thecodont hypothesis for the origin of birds is characterised by being a default option. This is because it is not due to positive correlation of characters and taxa but due to the negative association with other taxa. It was originally thought that theropods shared more features with birds than any other group. However, at the time there was no evidence for clavicles in theropods, which are the equivalent of the furcula or wishbone in birds. It was thought that clavicles could not have been lost and then re-evolved into furcula and so a more ancient ancestor for birds was sought for. The candidate suggested was the thecodonts from which all other archosaurs are thought to have evolved. A problem is that the archosaurs are a "wastebasket" group containing all archosaurian reptiles that do not fit into dinosaurs, crocodiles or pterosaurs. Hence, they have no diagnostic characters of their own and are not a "good" phylogenetic group. This makes it difficult to compare them with other taxonomic groups (Padian et al., 1998).
The Crocodylomorph Hypothesis
Crocodylomorphs include crocodiles and some Triassic-Jurassic forms that are closely related but not true crocodiles. This hypothesis has fewer problems than the thecodont hypothesis, as crocodiles are a monophyletic group, which can be compared to other taxa. The synapomorphies of birds and crocodiles have been tested, but it has been found that most are general to the archosaurs. Even if these are accepted, there are only 15-20 synapomorphies compared to over 70 with the theropods (Padian et al., 1998).
The Theropod Hypothesis
Cladistic analysis supports this hypothesis and shows that the most closely related group (sister group) of theropods to the birds includes the dromaesaurids such as Deinonychus. There are numerous synapomorphies of the skeleton and skull that link birds and theropods. There have been found a series of skeletal changes in theropods that are considered to be avian. For example, basal theropod dinosaurs have lightly built bones and a foot reduced to three main toes, with the first held off the ground and the fifth lost. Moving through the theropod sequence towards birds, there is a reduction and loss of manual digits four and five, increasing lightness of the skeleton, and a reduction in the number, and partial interlocking, of the tail vertebrae. In coelurosaurs, which include birds, dromaeosaurids and other groups, the arms become longer and the first toe begins to rotate backwards behind the metatarsals. Fused clavicles (furcula) are apparently basal to Tetanurae (carnosaurs and coelurosaurs) and sternal plates are known in a variety of tetanurans. In the pelvis, the pubis and ischium begin to show a greater disparity in length. Finally, in the dromaeosaurids and Archaeopteryx, the pubis begins to point backwards instead of forwards, the anterior projection on the foot of the pubis is lost, the tail becomes even shorter and the hyperflexing wrist joint is present, which allows the action that is crucial to the flight stroke in birds. These features were passed to birds from their dinosaurian ancestors and not specifically evolved for an avian lifestyle (Padian et al., 1998).